

Rare Phytocannabinoids Exert Anti-Inflammatory Effects on Human Keratinocytes via the Endocannabinoid System and MAPK Signaling Pathway
 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
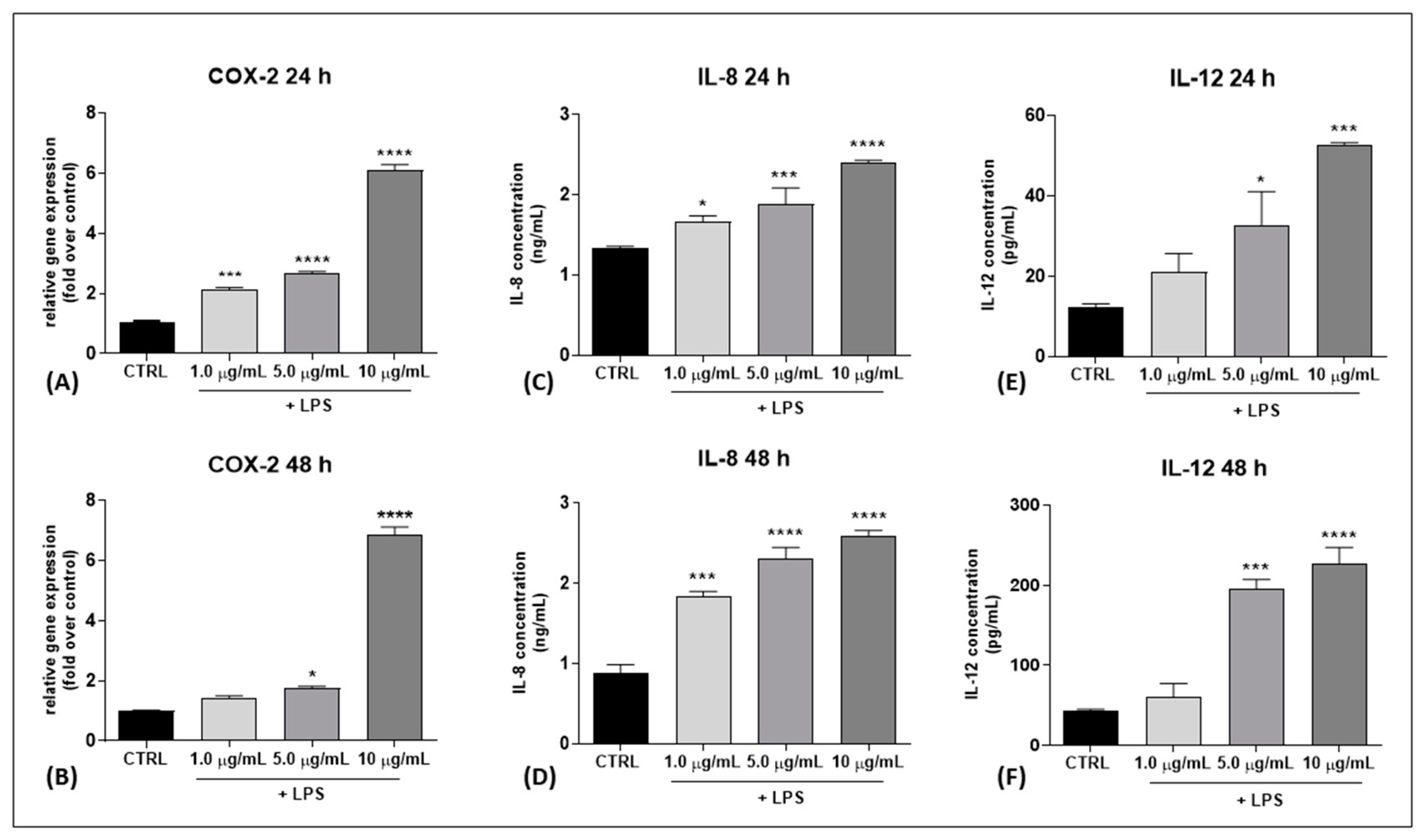
2.1. Set Up of an In Vitro Model of Inflamed Keratinocytes (HaCaT Cells)
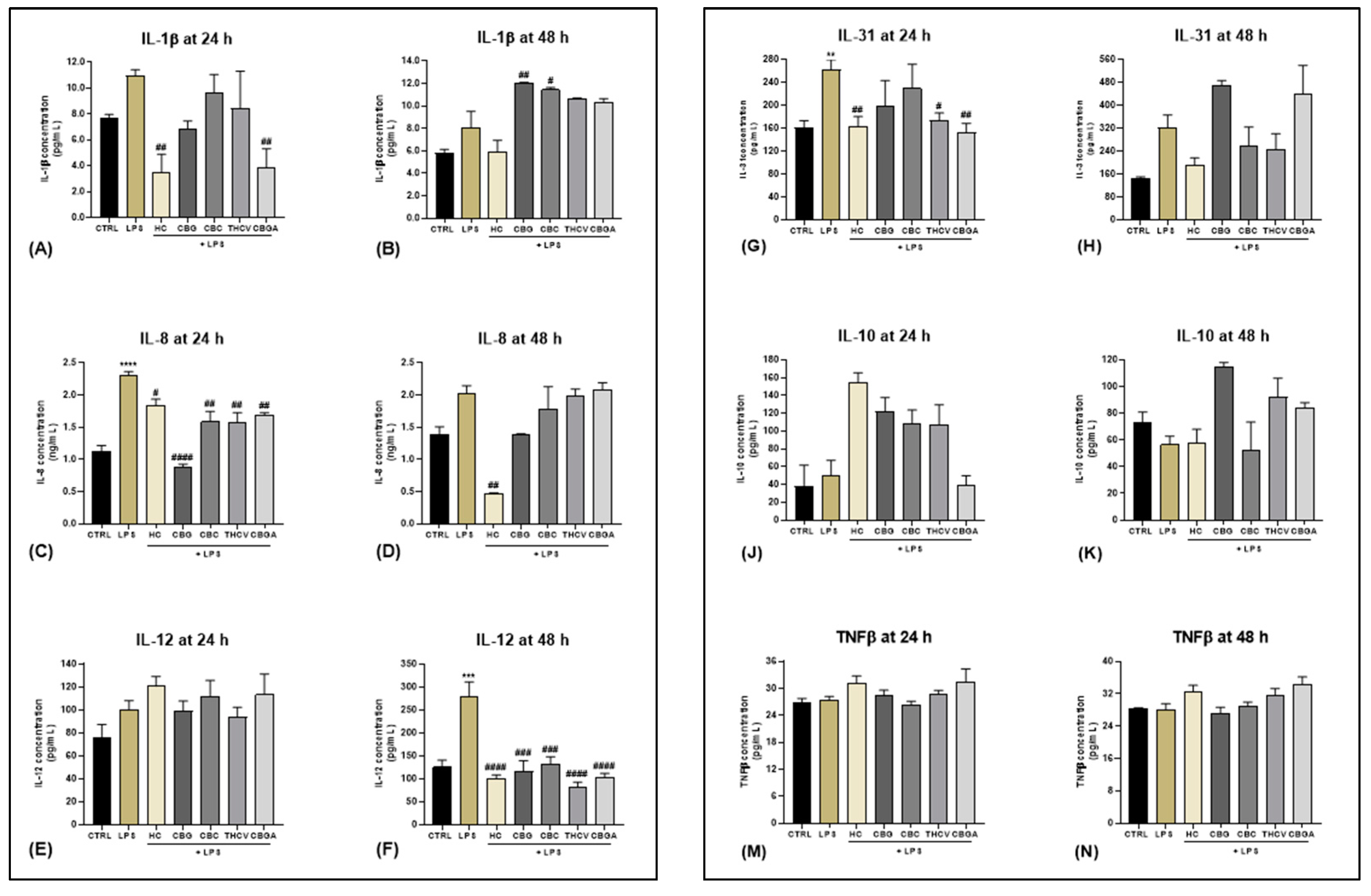
2.2. Effects of Rare pCBs on Cytokine Expression
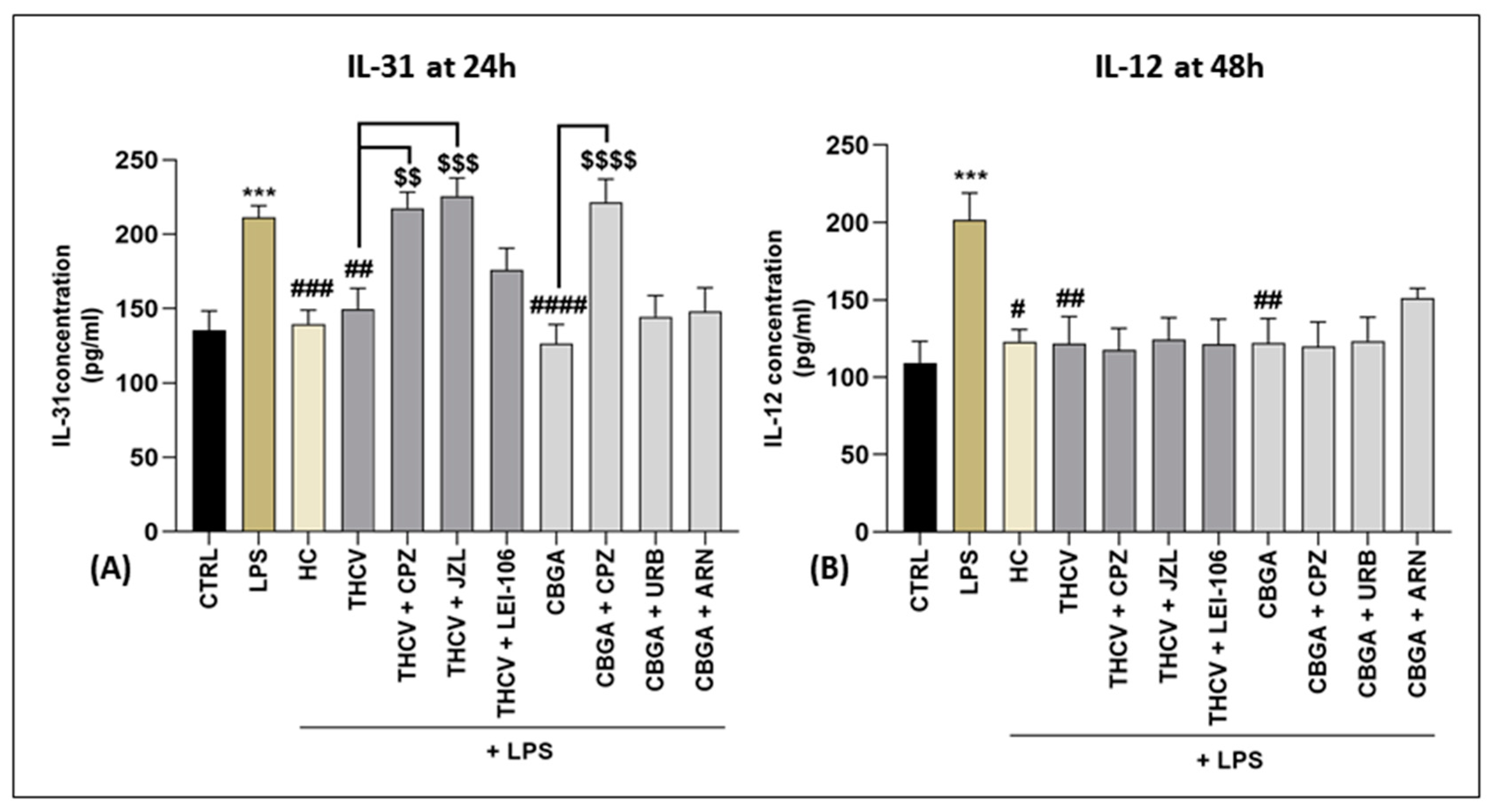
2.3. Role of the eCB System in Cytokine Release Mediated by THCV and CBGA
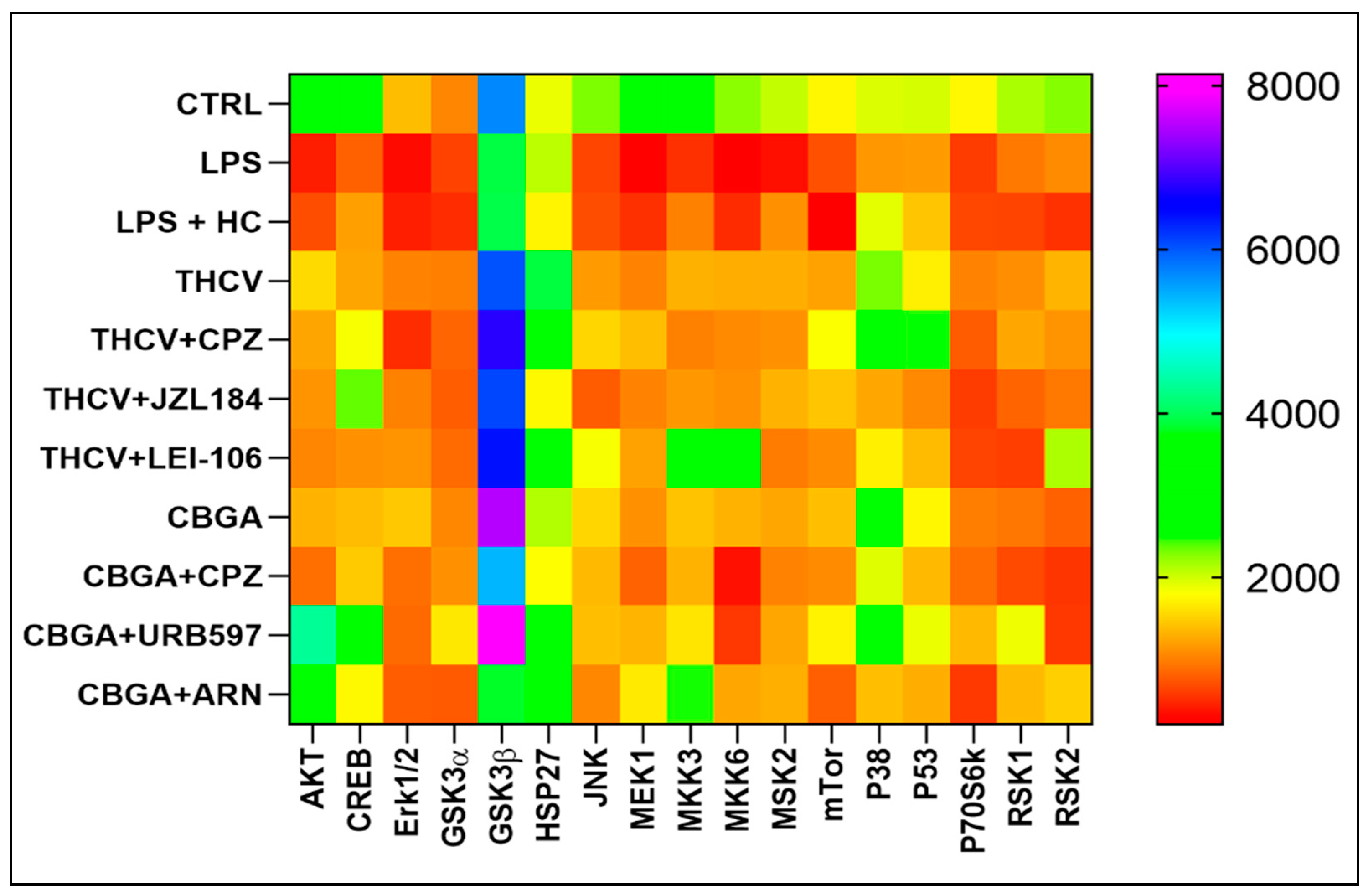
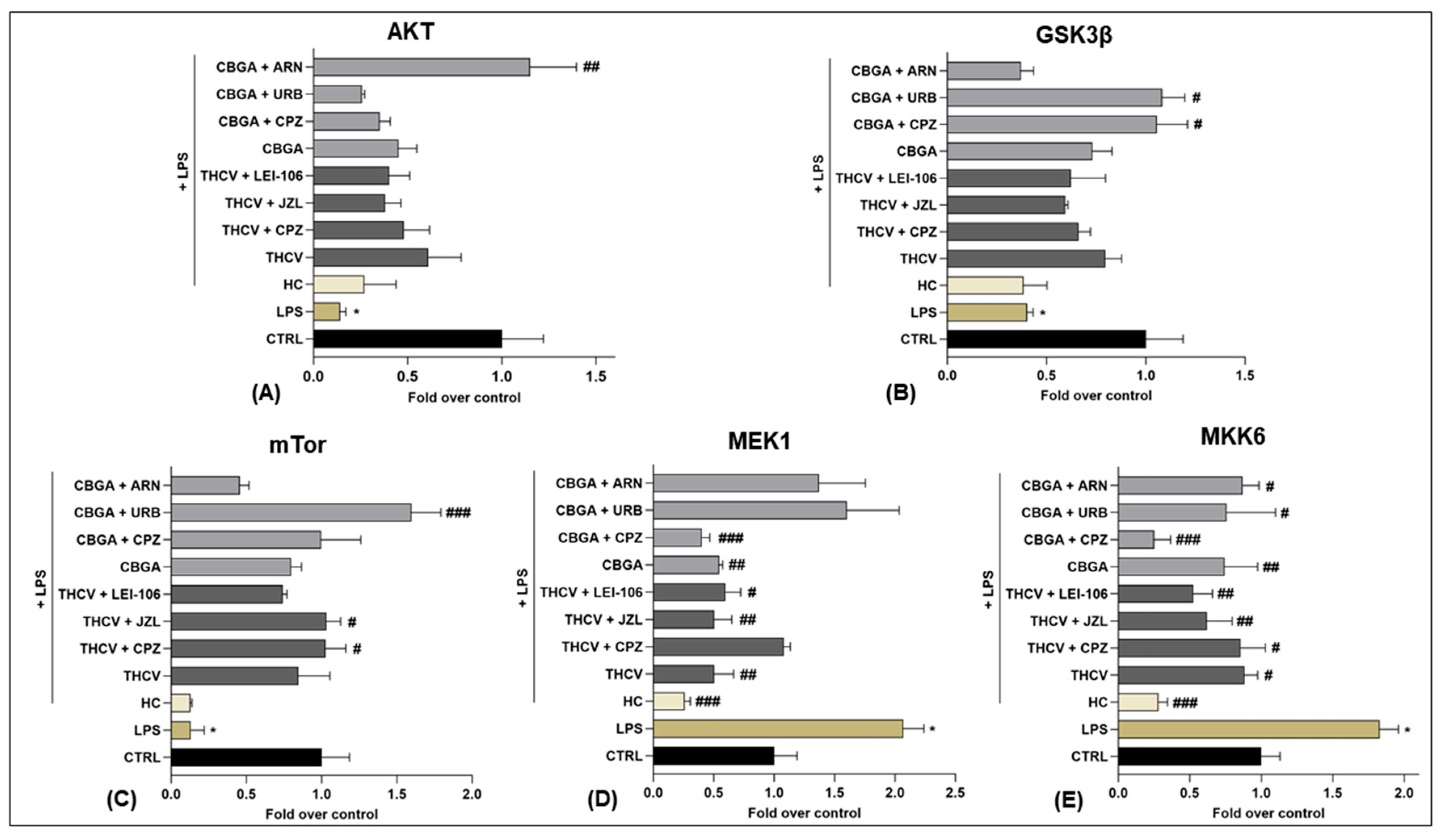
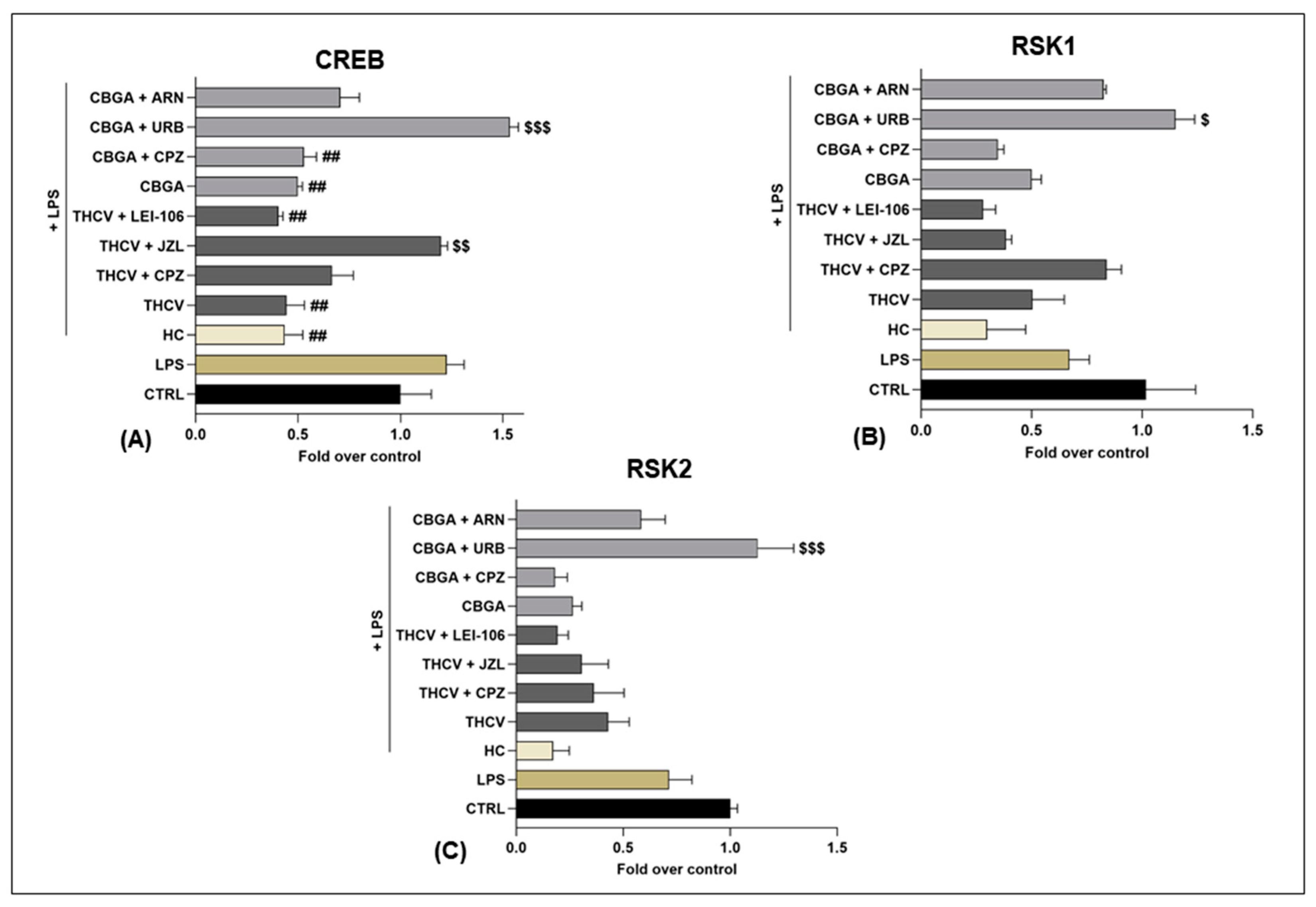
2.4. Effects of THCV and CBGA on the eCB-Dependent MAPK Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Line and Treatments
4.3. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. MAPK Signaling Pathway Array
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaid, N.A.M.; Sekar, M.; Bonam, S.R.; Gan, S.H.; Lum, P.T.; Begum, M.Y.; Rani, N.N.I.M.; Vaijanathappa, J.; Wu, Y.S.; Subramaniyan, V.; et al. Promising Natural Products in New Drug Design, Development, and Therapy for Skin Disorders: An Overview of Scientific Evidence and Understanding Their Mechanism of Action. Drug Des. Dev. Ther. 2022, 16, 23–66. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.M.; Gomes, A.L.; Boas, I.V.; Marto, J.; Ribeiro, H.M. Cannabis-Based Products for the Treatment of Skin Inflammatory Diseases: A Timely Review. Pharmaceuticals 2022, 15, 210. [Google Scholar] [CrossRef] [PubMed]
- Eagleston, L.R.M.; Kalani, N.K.; Patel, R.R.; Flaten, H.K.; Dunnick, C.A.; Dellavalle, R.P. Cannabinoids in Dermatology: A Scoping Review. Dermatol. Online J. 2018, 24. [Google Scholar] [CrossRef]
- Gülck, T.; Møller, B.L. Phytocannabinoids: Origins and Biosynthesis. Trends Plant Sci. 2020, 25, 985–1004. [Google Scholar] [CrossRef]
- ElSohly, M.A.; Radwan, M.M.; Gul, W.; Chandra, S.; Galal, A. Phytochemistry of Cannabis sativa L. Phytocannabinoids 2017, 103, 1–36. [Google Scholar] [CrossRef]
- Straiker, A.; Wilson, S.; Corey, W.; Dvorakova, M.; Bosquez, T.; Tracey, J.; Wilkowski, C.; Ho, K.; Wager-Miller, J.; Mackie, K. An Evaluation of Understudied Phytocannabinoids and Their Effects in Two Neuronal Models. Molecules 2021, 26, 5352. [Google Scholar] [CrossRef]
- Pattnaik, F.; Nanda, S.; Mohanty, S.; Dalai, A.K.; Kumar, V.; Ponnusamy, S.K.; Naik, S. Cannabis: Chemistry, Extraction and Therapeutic Applications. Chemosphere 2022, 289, 133012. [Google Scholar] [CrossRef]
- Di Meo, C.; Tortolani, D.; Standoli, S.; Angelucci, C.B.; Fanti, F.; Leuti, A.; Sergi, M.; Kadhim, S.; Hsu, E.; Rapino, C.; et al. Effects of Rare Phytocannabinoids on the Endocannabinoid System of Human Keratinocytes. Int. J. Mol. Sci. 2022, 23, 5430. [Google Scholar] [CrossRef]
- Bíró, T.; Tóth, B.I.; Haskó, G.; Paus, R.; Pacher, P. The Endocannabinoid System of the Skin in Health and Disease: Novel Perspectives and Therapeutic Opportunities. Trends Pharmacol. Sci. 2009, 30, 411–420. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bab, I.; Bíró, T.; Cabral, G.A.; Dey, S.K.; di Marzo, V.; Konje, J.C.; Kunos, G.; Mechoulam, R.; Pacher, P.; et al. Endocannabinoid Signaling at the Periphery: 50 Years after THC. Trends Pharmacol. Sci. 2015, 36, 277–296. [Google Scholar] [CrossRef] [Green Version]
- Baswan, S.M.; Klosner, A.E.; Glynn, K.; Rajgopal, A.; Malik, K.; Yim, S.; Stern, N. Therapeutic Potential of Cannabidiol (CBD) for Skin Health and Disorders. Clin. Cosmet. Investig. Dermatol. 2020, 13, 927–942. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Marzo, V. di Cannabinoids and the Expanded Endocannabinoid System in Neurological Disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Z.; Uyama, T.; Tsuboi, K.; Ueda, N. Mammalian Enzymes Responsible for the Biosynthesis of N-Acylethanolamines. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1546–1561. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M. Metabolism of the Endocannabinoid Anandamide: Open Questions after 25 Years. Front. Mol. Neurosci. 2017, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Baggelaar, M.P.; Maccarrone, M.; van der Stelt, M. 2-Arachidonoylglycerol: A Signaling Lipid with Manifold Actions in the Brain. Prog. Lipid Res. 2018, 71, 1–17. [Google Scholar] [CrossRef]
- Del Río, C.; Millán, E.; García, V.; Appendino, G.; DeMesa, J.; Muñoz, E. The Endocannabinoid System of the Skin. A Potential Approach for the Treatment of Skin Disorders. Biochem. Pharmacol. 2018, 157, 122–133. [Google Scholar] [CrossRef]
- Ramer, R.; Hinz, B. Cannabinoid Compounds as a Pharmacotherapeutic Option for the Treatment of Non-Cancer Skin Diseases. Cells 2022, 11, 4102. [Google Scholar] [CrossRef]
- Tóth, K.F.; Ádám, D.; Bíró, T.; Oláh, A. Cannabinoid Signaling in the Skin: Therapeutic Potential of the “c(Ut)Annabinoid” System. Molecules 2019, 24, 918. [Google Scholar] [CrossRef]
- Jiang, Y.; Tsoi, L.C.; Billi, A.C.; Ward, N.L.; Harms, P.W.; Zeng, C.; Maverakis, E.; Kahlenberg, J.M.; Gudjonsson, J.E. Cytokinocytes: The Diverse Contribution of Keratinocytes to Immune Responses in Skin. JCI Insight 2020, 5, e142067. [Google Scholar] [CrossRef]
- Piipponen, M.; Li, D.; Landén, N.X. The Immune Functions of Keratinocytes in Skin Wound Healing. Int. J. Mol. Sci. 2020, 21, 8790. [Google Scholar] [CrossRef]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin Microbiota–Host Interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in Wound Healing: A Comprehensive Review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiù, V.; Rapino, C.; Talamonti, E.; Leuti, A.; Lanuti, M.; Gueniche, A.; Jourdain, R.; Breton, L.; Maccarrone, M. Anandamide Suppresses Proinflammatory T Cell Responses In Vitro through Type-1 Cannabinoid Receptor–Mediated MTOR Inhibition in Human Keratinocytes. J. Immunol. 2016, 197, 3545–3553. [Google Scholar] [CrossRef] [PubMed]
- Pasquariello, N.; Oddi, S.; Malaponti, M.; Maccarrone, M. Chapter 17 Regulation of Gene Transcription and Keratinocyte Differentiation by Anandamide. Vitam Horm. 2009, 81, 441–467. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.L.; Mukhtar, H.; Bickers, D.R.; Kopelovich, L.; Athar, M. Cyclooxygenases in the Skin: Pharmacological and Toxicological Implications. Toxicol. Appl. Pharmacol. 2003, 192, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Hyun, E.; Bolla, M.; Steinhoff, M.; Wallace, J.L.; del Soldato, P.; Vergnolle, N. Anti-Inflammatory Effects of Nitric Oxide-Releasing Hydrocortisone NCX 1022, in a Murine Model of Contact Dermatitis. Br. J. Pharmacol. 2004, 143, 618–625. [Google Scholar] [CrossRef]
- Tóth, B.I.; Dobrosi, N.; Dajnoki, A.; Czifra, G.; Oláh, A.; Szöllősi, A.G.; Juhász, I.; Sugawara, K.; Paus, R.; Bíró, T. Endocannabinoids Modulate Human Epidermal Keratinocyte Proliferation and Survival via the Sequential Engagement of Cannabinoid Receptor-1 and Transient Receptor Potential Vanilloid-1. J. Investig. Dermatol. 2011, 131, 1095–1104. [Google Scholar] [CrossRef]
- Van der Wel, T.; Janssen, F.J.; Baggelaar, M.P.; Deng, H.; den Dulk, H.; Overkleeft, H.S.; van der Stelt, M. A Natural Substrate-Based Fluorescence Assay for Inhibitor Screening on Diacylglycerol Lipase α. J. Lipid Res. 2015, 56, 927–935. [Google Scholar] [CrossRef]
- Tanaka, M.; Yagyu, K.; Sackett, S.; Zhang, Y. Anti-Inflammatory Effects by Pharmacological Inhibition or Knockdown of Fatty Acid Amide Hydrolase in BV2 Microglial Cells. Cells 2019, 8, 491. [Google Scholar] [CrossRef]
- Castellani, B.; Diamanti, E.; Pizzirani, D.; Tardia, P.; Maccesi, M.; Realini, N.; Magotti, P.; Garau, G.; Bakkum, T.; Rivara, S.; et al. Synthesis and Characterization of the First Inhibitor of N -Acylphosphatidylethanolamine Phospholipase D (NAPE-PLD). Chem. Commun. 2017, 53, 12814–12817. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-J.; Song, E.-A.C.; Jung, M.-S.; Choi, S.-H.; Baik, H.-H.; Jin, B.K.; Kim, J.H.; Chung, S.-H. Phosphorylation and Inactivation of Glycogen Synthase Kinase 3β (GSK3β) by Dual-Specificity Tyrosine Phosphorylation-Regulated Kinase 1A (Dyrk1A). J. Biol. Chem. 2015, 290, 2321–2333. [Google Scholar] [CrossRef] [PubMed]
- Frame, S.; Cohen, P. GSK3 Takes Centre Stage More than 20 Years after Its Discovery. Biochem. J. 2001, 359, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis Sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Pirazzi, V.; Pasquariello, N.; Maccarrone, M. Endocannabinoid Signaling and Epidermal Differentiation. Eur. J. Dermatol. 2011, 21, 29–34. [Google Scholar] [CrossRef]
- Müller, G.; Saloga, J.; Germann, T.; Bellinghausen, I.; Mohamadzadeh, M.; Knop, J.; Enk, A.H. Identification and Induction of Human Keratinocyte-Derived IL-12. J. Clin. Investig. 1994, 94, 1799–1805. [Google Scholar] [CrossRef]
- Murata, S.; Kaneko, S.; Morita, E. Interleukin-8 Levels in the Stratum Corneum as a Biomarker for Monitoring Therapeutic Effect in Atopic Dermatitis Patients. Int. Arch. Allergy Immunol. 2021, 182, 592–606. [Google Scholar] [CrossRef]
- Yawalkar, N.; Egli, F.; Brand, C.U.; Pichler, W.J.; Braathen, L.R. Antigen-Presenting Cells and Keratinocytes Express Interleukin-12 in Allergic Contact Dermatitis. Contact Dermat. 2000, 42, 18–22. [Google Scholar] [CrossRef]
- Borgia, F.; Custurone, P.; Pomi, F.L.; Cordiano, R.; Alessandrello, C.; Gangemi, S. IL-31: State of the Art for an Inflammation-Oriented Interleukin. Int. J. Mol. Sci. 2022, 23, 6507. [Google Scholar] [CrossRef]
- Gibbs, B.F.; Patsinakidis, N.; Raap, U. Role of the Pruritic Cytokine IL-31 in Autoimmune Skin Diseases. Front. Immunol. 2019, 10, 1383. [Google Scholar] [CrossRef]
- Kulig, P.; Musiol, S.; Freiberger, S.N.; Schreiner, B.; Gyülveszi, G.; Russo, G.; Pantelyushin, S.; Kishihara, K.; Alessandrini, F.; Kündig, T.; et al. IL-12 Protects from Psoriasiform Skin Inflammation. Nat. Commun. 2016, 7, 13466. [Google Scholar] [CrossRef] [PubMed]
- Oláh, A.; Markovics, A.; Szabó-Papp, J.; Szabó, P.T.; Stott, C.; Zouboulis, C.C.; Bíró, T. Differential Effectiveness of Selected Non-Psychotropic Phytocannabinoids on Human Sebocyte Functions Implicates Their Introduction in Dry/Seborrhoeic Skin and Acne Treatment. Exp. Dermatol. 2016, 25, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Nattkemper, L.A.; Tey, H.L.; Valdes-Rodriguez, R.; Lee, H.; Mollanazar, N.K.; Albornoz, C.; Sanders, K.M.; Yosipovitch, G. The Genetics of Chronic Itch: Gene Expression in the Skin of Patients with Atopic Dermatitis and Psoriasis with Severe Itch. J. Investig. Dermatol. 2018, 138, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Follansbee, T.; Wu, X.; Han, D.; Yu, S.; Domocos, D.T.; Shi, Z.; Carstens, M.; Carstens, E.; Hwang, S.T. TRPV1 Mediates Inflammation and Hyperplasia in Imiquimod (IMQ)-Induced Psoriasiform Dermatitis (PsD) in Mice. J. Dermatol. Sci. 2018, 92, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yang, P.; Mack, M.R.; Dryn, D.; Luo, J.; Gong, X.; Liu, S.; Oetjen, L.K.; Zholos, A.V.; Mei, Z.; et al. Sensory TRP Channels Contribute Differentially to Skin Inflammation and Persistent Itch. Nat. Commun. 2017, 8, 980. [Google Scholar] [CrossRef] [PubMed]
- Yesilyurt, O.; Cayirli, M.; Sakin, Y.S.; Seyrek, M.; Akar, A.; Dogrul, A. Systemic and Spinal Administration of FAAH, MAGL Inhibitors and Dual FAAH/MAGL Inhibitors Produce Antipruritic Effect in Mice. Arch. Dermatol. Res. 2016, 308, 335–345. [Google Scholar] [CrossRef]
- Haruna, T.; Soga, M.; Morioka, Y.; Imura, K.; Furue, Y.; Yamamoto, M.; Hayakawa, J.; Deguchi, M.; Arimura, A.; Yasui, K. The Inhibitory Effect of S-777469, a Cannabinoid Type 2 Receptor Agonist, on Skin Inflammation in Mice. Pharmacology 2017, 99, 259–267. [Google Scholar] [CrossRef]
- Hoffmeister, L.; Diekmann, M.; Brand, K.; Huber, R. GSK3: A Kinase Balancing Promotion and Resolution of Inflammation. Cells 2020, 9, 820. [Google Scholar] [CrossRef]
- Kaidanovich-Beilin, O.; Woodgett, J.R. GSK-3: Functional Insights from Cell Biology and Animal Models. Front. Mol. Neurosci. 2011, 4, 40. [Google Scholar] [CrossRef]
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The Role of the Transcription Factor CREB in Immune Function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef] [Green Version]
- Bansal, M.; Singh, N.; Alam, S.; Pal, S.; Satyanarayana, G.N.V.; Singh, D.; Ansari, K.M. Alternariol Induced Proliferation in Primary Mouse Keratinocytes and Inflammation in Mouse Skin Is Regulated via PGE2/EP2/CAMP/p-CREB Signaling Pathway. Toxicology 2019, 412, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Funding, A.T.; Johansen, C.; Kragballe, K.; Otkjær, K.; Jensen, U.B.; Madsen, M.W.; Fjording, M.S.; Finnemann, J.; Skak-Nielsen, T.; Paludan, S.R.; et al. Mitogen- and Stress-Activated Protein Kinase 1 Is Activated in Lesional Psoriatic Epidermis and Regulates the Expression of Pro-Inflammatory Cytokines. J. Investig. Dermatol. 2006, 126, 1784–1791. [Google Scholar] [CrossRef]
- Funding, A.T.; Johansen, C.; Kragballe, K.; Iversen, L. Mitogen- and Stress-Activated Protein Kinase 2 and Cyclic AMP Response Element Binding Protein Are Activated in Lesional Psoriatic Epidermis. J. Investig. Dermatol. 2007, 127, 2012–2019. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.C.; Antoniades, W.; Okalova, J.; Roos, M.M.; Grimsey, N.J. Atypical P38 Signaling, Activation, and Implications for Disease. Int. J. Mol. Sci. 2021, 22, 4183. [Google Scholar] [CrossRef]
- Che, G.; Wang, Y.; Zhou, B.; Gao, L.; Wang, T.; Yuan, F.; Zhang, L. Knockdown of Heparanase Suppresses Invasion of Human Trophoblasts by Activating P38 MAPK Signaling Pathway. Dis. Markers 2018, 2018, 7413027. [Google Scholar] [CrossRef] [PubMed]
- Lyon, E. Mutation Detection Using Fluorescent Hybridization Probes and Melting Curve Analysis. Expert Rev. Mol. Diagn. 2001, 1, 92–101. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- de Camargo, R.W.; de Novais Júnior, L.R.; da Silva, L.M.; Meneguzzo, V.; Daros, G.C.; da Silva, M.G.; de Bitencourt, R.M. Implications of the Endocannabinoid System and the Therapeutic Action of Cannabinoids in Autism Spectrum Disorder: A Literature Review. Pharmacol. Biochem. Behav. 2022, 221, 173492. [Google Scholar] [CrossRef]
- Lah, T.T.; Majc, B.; Novak, M.; Sušnik, A.; Breznik, B.; Porčnik, A.; Bošnjak, R.; Sadikov, A.; Malavolta, M.; Halilčević, S.; et al. The Cytotoxic Effects of Cannabidiol and Cannabigerol on Glioblastoma Stem Cells May Mostly Involve GPR55 and TRPV1 Signaling. Cancers 2022, 14, 5918. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CBG | CBC | THCV | CBGA | |||||
|---|---|---|---|---|---|---|---|---|
| Interleukins | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h |
| IL-1β | ↑ ++ | ↑ + | ↓ ++ | |||||
| IL-8 | ↓ ++++ | ↓ ++ | ↓ ++ | ↓ ++ | ||||
| IL-12 | ↓ +++ | ↓ +++ | ↓ ++++ | ↓ ++++ | ||||
| IL-31 | ↓ + | ↓ ++ | ||||||
| THCV Effects | CBGA Effects | |||||||
|---|---|---|---|---|---|---|---|---|
| MAPKs | THCV | THCV + TRPV1 Antagonist (CPZ) | THCV + MAGL Inhibitor (JZL184) | THCV + DAGLs Inhibitor (LEI-106) | CBGA | CBGA + TRPV1 Antagonist (CPZ) | CBGA + FAAH Inhibitor (URB597) | CBGA + NAPE-PLD Inhibitor (ARN19874) |
| AKT | ↑ ++ vs. LPS | |||||||
| GSK3β | ↑ + vs. LPS | ↑ + vs. LPS | ||||||
| mTor | ↑ + vs. LPS | ↑ + vs. LPS | ↑ +++ vs. LPS | |||||
| MEK1 | ↓ ++ vs. LPS | ↓ ++ vs. LPS | ↓ + vs. LPS | ↓ ++ vs. LPS | ↓ +++ vs. LPS | |||
| MKK6 | ↓ + vs. LPS | ↓ + vs. LPS | ↓ ++ vs. LPS | ↓ ++ vs. LPS | ↓ ++ vs. LPS | ↓ +++ vs. LPS | ↓ + vs. LPS | ↓ + vs. LPS |
| CREB | ↓ ++ vs. LPS | ↑ ++ vs. THCV | ↓ ++ vs. LPS | ↓ ++ vs. LPS | ↓ ++ vs. LPS | ↑ +++ vs. CBGA | ||
| RSK1 | ↑ + vs. CBGA | |||||||
| RSK2 | ↑ +++ vs. CBGA | |||||||
| Gene | Forward Primer Sequence (5′→3′) | Reverse Primer Sequence (5′→3′) |
|---|---|---|
| PTGS2 | ATACTTACCCACTTCAAGGG | ATCAGGAAGCTGCTTTTTACC |
| ACTB | TGACCCAGATCATGTTTGAG | TTAATGTCACGCACGATTTCC |
| GAPDH | CAGCCTCAAGATCATCAGCA | TGTGGTCATGAGTCCTTCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tortolani, D.; Di Meo, C.; Standoli, S.; Ciaramellano, F.; Kadhim, S.; Hsu, E.; Rapino, C.; Maccarrone, M. Rare Phytocannabinoids Exert Anti-Inflammatory Effects on Human Keratinocytes via the Endocannabinoid System and MAPK Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 2721. https://doi.org/10.3390/ijms24032721
Tortolani D, Di Meo C, Standoli S, Ciaramellano F, Kadhim S, Hsu E, Rapino C, Maccarrone M. Rare Phytocannabinoids Exert Anti-Inflammatory Effects on Human Keratinocytes via the Endocannabinoid System and MAPK Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(3):2721. https://doi.org/10.3390/ijms24032721
Chicago/Turabian StyleTortolani, Daniel, Camilla Di Meo, Sara Standoli, Francesca Ciaramellano, Salam Kadhim, Eric Hsu, Cinzia Rapino, and Mauro Maccarrone. 2023. "Rare Phytocannabinoids Exert Anti-Inflammatory Effects on Human Keratinocytes via the Endocannabinoid System and MAPK Signaling Pathway" International Journal of Molecular Sciences 24, no. 3: 2721. https://doi.org/10.3390/ijms24032721
APA StyleTortolani, D., Di Meo, C., Standoli, S., Ciaramellano, F., Kadhim, S., Hsu, E., Rapino, C., & Maccarrone, M. (2023). Rare Phytocannabinoids Exert Anti-Inflammatory Effects on Human Keratinocytes via the Endocannabinoid System and MAPK Signaling Pathway. International Journal of Molecular Sciences, 24(3), 2721. https://doi.org/10.3390/ijms24032721